
Introducción
El canal TRPM8 (Transient Receptor Potential Melastatin 8) es una proteína transmembrana perteneciente a la superfamilia de canales iónicos TRP, implicada en la detección y transducción de estímulos sensoriales en organismos vertebrados. Dentro de esta familia, TRPM8 se clasifica en la subfamilia melastatina (TRPM) y se reconoce como uno de los principales sensores moleculares responsables de la percepción del frío ambiental y químico (McKemy et al., 2002; Peier et al., 2002).
TRPM8 fue identificado a principios del siglo XXI en el contexto de estudios orientados a desentrañar los mecanismos moleculares de la termorrecepción. Su descubrimiento representó un avance clave en neurobiología sensorial al establecer una base molecular para la detección de temperaturas frías, en paralelo a otros canales TRP implicados en la percepción térmica, como TRPV1 en la detección de calor nocivo (Caterina et al., 1997; McKemy et al., 2002).
Desde el punto de vista funcional, TRPM8 actúa como un canal catiónico no selectivo activado por descensos de temperatura —generalmente en el rango inferior a 25–28 °C— así como por compuestos químicos como el mentol, responsable de la sensación refrescante característica en piel y mucosas. Su activación permite el flujo de iones como calcio (Ca²⁺) y sodio (Na⁺), lo que induce despolarización neuronal y la generación de señales eléctricas interpretadas como frío por el sistema nervioso central (Peier et al., 2002; Bautista et al., 2007).
A nivel de expresión tisular, TRPM8 se localiza predominantemente en neuronas sensoriales periféricas de pequeño diámetro, especialmente en los ganglios de la raíz dorsal y ganglios trigeminales. Estas neuronas forman parte de las fibras aferentes primarias encargadas de transmitir información térmica desde la periferia hacia el sistema nervioso central, constituyendo un componente esencial del sistema somatosensorial (Dhaka et al., 2007; Knowlton et al., 2010).
Más allá de su papel clásico en la termorrecepción, estudios posteriores han descrito la expresión de TRPM8 en tejidos no neuronales, incluyendo epitelios y próstata, lo que sugiere funciones adicionales en procesos fisiológicos periféricos que aún están en fase de caracterización (Tsavaler et al., 2001; Bidaux et al., 2007).
En términos biofísicos, TRPM8 es un canal cuya actividad está regulada no solo por estímulos térmicos y químicos, sino también por el entorno lipídico de la membrana, especialmente por fosfolípidos como el fosfatidilinositol 4,5-bisfosfato (PIP2). Esta dependencia introduce un nivel de regulación que conecta su activación con rutas de señalización intracelular más amplias (Rohács et al., 2005; Liu & Qin, 2005).
Desde una perspectiva integradora, TRPM8 no debe considerarse únicamente como un sensor de frío, sino como una diana molecular multifuncional cuya actividad puede ser modulada por distintos mediadores endógenos y exógenos. Entre estos se incluyen lípidos bioactivos y compuestos de origen vegetal, lo que amplía su relevancia en fisiología sensorial y farmacología (Bandell et al., 2004; Vriens et al., 2014).
En este contexto, TRPM8 adquiere interés dentro del estudio del sistema endocannabinoide y sus dianas moleculares asociadas, ya que diversos fitocannabinoides y lípidos endógenos han demostrado capacidad para modular canales TRP. Estas interacciones sitúan a TRPM8 en la interfaz entre señalización sensorial, regulación lipídica y farmacología de cannabinoides (De Petrocellis et al., 2011; Muller et al., 2019).
El estudio de TRPM8 continúa siendo un área activa de investigación tanto en neurociencia básica como en contextos aplicados relacionados con la percepción sensorial y la modulación de señales periféricas. Su caracterización ha contribuido de manera significativa a la comprensión molecular de la termorrecepción, consolidándolo como un componente fundamental del sistema sensorial (Dhaka et al., 2006; Vriens et al., 2014).
Historia del descubrimiento
La identificación de TRPM8 se produjo a comienzos de la década de 2000 en el contexto de una intensa búsqueda por comprender los mecanismos moleculares responsables de la termorrecepción. Hasta ese momento, aunque se conocían respuestas neuronales al frío, no se había identificado de forma clara el receptor específico encargado de detectar temperaturas bajas a nivel molecular (McKemy et al., 2002; Peier et al., 2002).
El descubrimiento de TRPM8 se llevó a cabo de forma independiente por dos grupos de investigación que emplearon estrategias de clonación funcional basadas en la respuesta celular a compuestos químicos conocidos por inducir sensación de frío, especialmente el mentol. Este enfoque permitió identificar un canal iónico cuya activación reproducía las características fisiológicas de las neuronas sensibles al frío (McKemy et al., 2002).
En el estudio de McKemy y colaboradores, se utilizó mentol como herramienta farmacológica para seleccionar células que expresaban proteínas sensibles a este compuesto. A través de este proceso, se logró aislar un canal previamente no caracterizado que respondía tanto al mentol como a descensos de temperatura, estableciendo una conexión directa entre estímulo químico y térmico en la activación del mismo receptor (McKemy et al., 2002).
De manera paralela, el grupo de Peier et al. identificó el mismo canal mediante un enfoque complementario, confirmando que su expresión en sistemas celulares heterólogos confería sensibilidad tanto al frío como a agentes refrescantes. Este hallazgo reforzó la idea de que TRPM8 actuaba como un detector molecular primario de estímulos fríos (Peier et al., 2002).
Estos trabajos se produjeron pocos años después de la caracterización de TRPV1 como receptor del calor y la capsaicina, lo que contribuyó a consolidar el concepto de que diferentes canales TRP funcionan como sensores térmicos especializados. En este contexto, TRPM8 pasó a representar el componente frío del sistema, completando el mapa inicial de la termorrecepción molecular (Caterina et al., 1997; Dhaka et al., 2006).
Tras su identificación, estudios posteriores se centraron en caracterizar su expresión en neuronas sensoriales periféricas, confirmando su localización en subpoblaciones específicas de fibras aferentes primarias. Estos trabajos demostraron que la activación de TRPM8 en dichas neuronas era suficiente para generar respuestas conductuales asociadas a la percepción de frío en modelos animales (Dhaka et al., 2007).
Además, el uso de modelos knockout para TRPM8 permitió validar su papel funcional, ya que los animales deficientes en este canal mostraban una reducción significativa en la capacidad de detectar estímulos fríos, lo que proporcionó evidencia directa de su relevancia fisiológica en la termorrecepción (Bautista et al., 2007; Dhaka et al., 2007).
En conjunto, el descubrimiento de TRPM8 no solo identificó el receptor molecular del frío, sino que también consolidó un cambio de paradigma en neurobiología sensorial, estableciendo que la percepción térmica se basa en la activación de canales iónicos específicos sensibles a rangos concretos de temperatura. Este avance sentó las bases para la exploración de nuevas dianas moleculares implicadas en la percepción sensorial y su modulación farmacológica (Vriens et al., 2014).
Estructura y arquitectura del canal TRPM8
TRPM8 es un canal iónico transmembrana perteneciente a la familia TRP, caracterizado por una arquitectura típica de canales dependientes de voltaje modificados. A nivel estructural, está formado por cuatro subunidades (homotetrámero), cada una de las cuales contribuye a la formación del poro central a través del cual se produce el flujo de iones (Yin et al., 2018; Diver et al., 2019).
Cada subunidad de TRPM8 contiene seis dominios transmembrana (S1–S6), organizados de manera similar a los canales de potasio dependientes de voltaje. Entre los segmentos S5 y S6 se localiza la región del poro, responsable de la permeabilidad iónica y la selectividad del canal, permitiendo el paso de cationes como Ca²⁺ y Na⁺ tras su activación (Voets et al., 2004; Yin et al., 2018).
El dominio S1–S4 actúa como región sensora, implicada en la detección de cambios en el entorno físico y químico. Aunque TRPM8 no es un canal dependiente de voltaje en el sentido clásico, esta región participa en la transducción de señales térmicas y en la interacción con agonistas como el mentol, facilitando los cambios conformacionales necesarios para la apertura del canal (Voets et al., 2004; Bandell et al., 2004).
En la porción intracelular N-terminal, TRPM8 presenta dominios característicos de la familia TRPM, incluyendo regiones ricas en repeticiones tipo ankyrin (ARDs), que desempeñan un papel clave en la estabilidad estructural del canal y en la interacción con proteínas reguladoras y componentes del citoesqueleto (Erler et al., 2006; Yin et al., 2018).
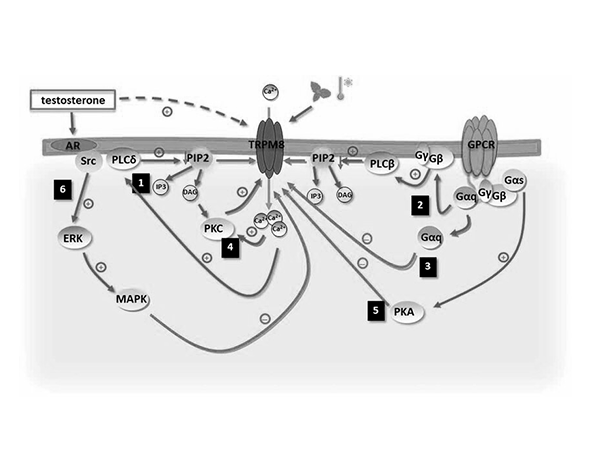
Por su parte, el extremo C-terminal contiene dominios implicados en la regulación funcional del canal, incluyendo regiones de unión a lípidos de membrana como el fosfatidilinositol 4,5-bisfosfato (PIP2), esencial para la actividad basal y la sensibilidad del canal. La interacción con PIP2 modula la probabilidad de apertura del canal y su respuesta a estímulos térmicos y químicos (Rohács et al., 2005; Liu & Qin, 2005).
Estudios estructurales recientes mediante criomicroscopía electrónica (cryo-EM) han permitido visualizar TRPM8 en distintos estados conformacionales, revelando cambios estructurales asociados a su activación. Estos trabajos han mostrado cómo la unión de agonistas y la disminución de temperatura inducen reorganizaciones en el dominio transmembrana que culminan en la apertura del poro iónico (Yin et al., 2018; Diver et al., 2019).
A nivel funcional, la arquitectura de TRPM8 permite integrar múltiples señales —térmicas, químicas y lipídicas— en un único evento de apertura del canal, lo que lo convierte en una plataforma molecular altamente sensible y modulable. Esta capacidad de integración es clave para su papel en la detección precisa de estímulos ambientales (Vriens et al., 2014).
En conjunto, la estructura de TRPM8 refleja un diseño molecular adaptado a la detección de frío y a la modulación por ligandos exógenos y endógenos, combinando elementos típicos de canales iónicos con dominios especializados que permiten su regulación fina dentro del entorno celular (Yin et al., 2018).
Distribución y expresión tisular
La expresión de TRPM8 se localiza principalmente en el sistema nervioso periférico, donde se encuentra en subpoblaciones específicas de neuronas sensoriales implicadas en la detección de estímulos térmicos. En particular, TRPM8 se expresa en neuronas de pequeño diámetro de los ganglios de la raíz dorsal (DRG) y de los ganglios trigeminales, que forman parte de las fibras aferentes primarias responsables de transmitir información sensorial desde la periferia hacia el sistema nervioso central (McKemy et al., 2002; Dhaka et al., 2007).
Estas neuronas TRPM8-positivas suelen corresponder a fibras de tipo C y Aδ, especializadas en la detección de estímulos térmicos no nocivos y, en determinados contextos, estímulos fríos potencialmente nocivos. La activación de estas fibras permite codificar la intensidad del estímulo térmico y generar respuestas adaptativas frente a cambios en la temperatura ambiental (Knowlton et al., 2010; Vriens et al., 2014).
A nivel periférico, las terminaciones nerviosas que expresan TRPM8 se distribuyen en tejidos expuestos al entorno externo, como la piel y las mucosas. En la piel, estas terminaciones se encuentran en la epidermis y dermis superficial, donde actúan como sensores directos de temperatura. En el sistema trigeminal, TRPM8 participa en la detección de estímulos térmicos en la cavidad oral y nasal, contribuyendo a la sensación refrescante inducida por compuestos como el mentol (Dhaka et al., 2007; Bautista et al., 2007).
Además de su expresión neuronal, diversos estudios han identificado la presencia de TRPM8 en tejidos no neuronales. Entre ellos destacan epitelios, células del tracto respiratorio y tejido prostático, donde su función no está completamente definida pero se ha asociado a procesos de señalización celular, regulación del tono tisular y posibles roles en fisiología local (Tsavaler et al., 2001; Bidaux et al., 2007).
En el contexto del sistema urogenital, la expresión de TRPM8 en próstata ha sido objeto de especial interés, ya que se ha observado una regulación diferencial de su expresión en condiciones fisiológicas y patológicas. Aunque estos hallazgos han impulsado investigaciones en este ámbito, su papel funcional exacto sigue siendo objeto de estudio (Bidaux et al., 2007).
A nivel molecular, la distribución de TRPM8 no es homogénea, sino que está restringida a subpoblaciones neuronales específicas, lo que sugiere una especialización funcional dentro del sistema somatosensorial. Esta organización permite que distintas poblaciones de neuronas respondan a rangos térmicos concretos, contribuyendo a una codificación precisa de la temperatura (Dhaka et al., 2006; Knowlton et al., 2010).
Además, la expresión de TRPM8 puede ser modulada por factores fisiológicos y patológicos, incluyendo procesos inflamatorios y cambios en el microentorno tisular. Estas modificaciones en su expresión o sensibilidad pueden alterar la percepción del frío, contribuyendo a fenómenos como la hipersensibilidad al frío observada en ciertos estados patológicos (Vriens et al., 2014).
En conjunto, la distribución de TRPM8 refleja su papel central como sensor periférico de temperatura, con una organización anatómica y celular altamente especializada que permite integrar estímulos térmicos del entorno y traducirlos en señales neuronales específicas dentro del sistema somatosensorial (Dhaka et al., 2007).
Interacción con el sistema endocannabinoide
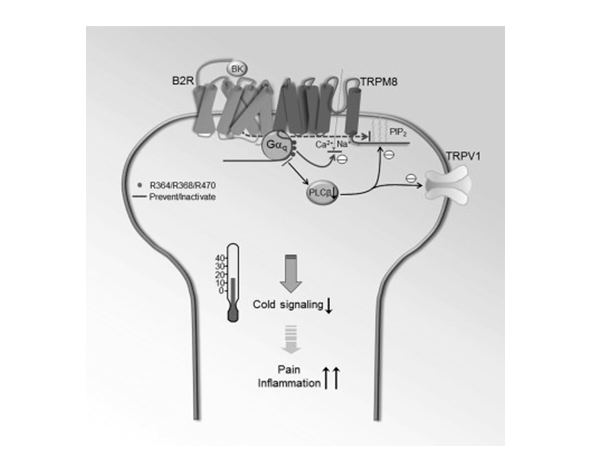
La relación entre TRPM8 y el sistema endocannabinoide (SEC) se enmarca dentro de un concepto más amplio de red de señalización lipídica, en la que endocannabinoides, fitocannabinoides y otros lípidos bioactivos modulan no solo receptores cannabinoides clásicos (CB1 y CB2), sino también un conjunto de dianas moleculares adicionales, entre las que se incluyen diversos canales TRP (Di Marzo et al., 2002; De Petrocellis & Di Marzo, 2009).
A diferencia de otros canales como TRPV1, donde la interacción con endocannabinoides como la anandamida está ampliamente caracterizada, la relación entre TRPM8 y el SEC se ha descrito principalmente en términos de modulación inhibitoria por ciertos ligandos cannabinoides. En este contexto, varios fitocannabinoides han mostrado capacidad para reducir la actividad de TRPM8, lo que sugiere un papel regulador indirecto sobre la señalización del frío (De Petrocellis et al., 2011).
Entre los compuestos estudiados, el cannabidiol (CBD) ha sido uno de los más investigados, observándose que puede inhibir la activación de TRPM8 en sistemas celulares experimentales. Este efecto no se produce a través de los receptores CB1 o CB2, sino mediante interacción directa o alostérica con el propio canal, lo que refuerza la idea de que los cannabinoides actúan sobre múltiples dianas más allá del sistema cannabinoide clásico (De Petrocellis et al., 2011; Muller et al., 2019).
Otros fitocannabinoides, como el Δ9-tetrahidrocannabinol (THC), también han mostrado capacidad para modular canales TRP, aunque su efecto sobre TRPM8 es más variable y dependiente del contexto experimental. En general, la evidencia sugiere que distintos cannabinoides pueden ejercer efectos diferenciales sobre estos canales, contribuyendo a perfiles farmacológicos complejos (De Petrocellis et al., 2011).
En el caso de los endocannabinoides, la evidencia sobre su interacción directa con TRPM8 es más limitada en comparación con TRPV1 o TRPA1. No obstante, ciertos lípidos endógenos estructuralmente relacionados, como los N-aciletanolamidas, pueden influir indirectamente en la actividad del canal a través de cambios en el entorno lipídico de la membrana o mediante la modulación de rutas intracelulares asociadas (Di Marzo et al., 2002; Vriens et al., 2014).
Además, la actividad de TRPM8 está estrechamente relacionada con la composición lipídica de la membrana, en particular con la disponibilidad de fosfolípidos como PIP2. Dado que el SEC participa en la regulación de lípidos de membrana y en la señalización intracelular asociada, se ha propuesto que puede influir indirectamente en la sensibilidad y activación de TRPM8 (Rohács et al., 2005; De Petrocellis & Di Marzo, 2009).
Desde una perspectiva funcional, esta interacción sitúa a TRPM8 dentro del denominado “endocannabinoidome”, un marco conceptual que amplía el SEC clásico para incluir múltiples mediadores lipídicos y dianas moleculares adicionales. En este sistema ampliado, los canales TRP actúan como integradores de señales químicas y físicas, contribuyendo a la regulación fina de la percepción sensorial (Di Marzo & Piscitelli, 2015).
En conjunto, aunque TRPM8 no forma parte del núcleo clásico del sistema endocannabinoide, su modulación por cannabinoides y otros lípidos bioactivos lo posiciona como una diana relevante en la interfaz entre señalización sensorial y regulación lipídica. Esta interacción contribuye a la complejidad funcional del SEC y refuerza la idea de que los efectos de los cannabinoides no pueden entenderse únicamente a través de CB1 y CB2 (De Petrocellis et al., 2011).
Mecanismos moleculares
El canal TRPM8 actúa como un sensor molecular polimodal cuya activación resulta de la integración de estímulos térmicos, químicos y lipídicos en una respuesta conformacional que culmina en la apertura del poro iónico. Este proceso implica una serie de cambios estructurales coordinados entre los dominios transmembrana y las regiones intracelulares del canal (Yin et al., 2018; Diver et al., 2019).
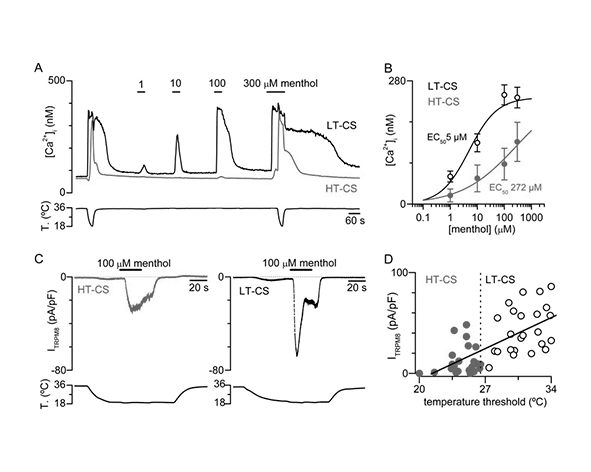
La activación por frío se produce a través de un mecanismo aún no completamente definido, pero que implica cambios en la dinámica de la membrana lipídica y en la estabilidad conformacional del canal. La disminución de temperatura favorece estados abiertos de TRPM8, probablemente mediante alteraciones en la interacción entre dominios transmembrana y el entorno lipídico que estabilizan la configuración conductora del canal (Voets et al., 2004; Vriens et al., 2014).
En paralelo, la activación química por agonistas como el mentol se basa en la unión directa de estas moléculas a sitios específicos dentro del dominio transmembrana. Esta interacción induce cambios conformacionales que mimetizan el efecto del frío, facilitando la apertura del canal incluso en ausencia de descensos térmicos significativos (Bandell et al., 2004; Yin et al., 2018).
Un elemento clave en la regulación de TRPM8 es su dependencia del fosfatidilinositol 4,5-bisfosfato (PIP2), un fosfolípido de membrana que actúa como cofactor esencial para su actividad. La depleción de PIP2, por ejemplo mediante activación de fosfolipasas, reduce significativamente la actividad del canal, lo que sugiere que este lípido estabiliza estados funcionales activos o facilita la transición hacia ellos (Rohács et al., 2005; Liu & Qin, 2005).
A nivel iónico, la apertura de TRPM8 permite la entrada de cationes, principalmente Ca²⁺ y Na⁺, generando despolarización de la membrana celular. El calcio intracelular no solo actúa como portador de carga, sino también como segundo mensajero que puede activar rutas de señalización adicionales, incluyendo mecanismos de retroalimentación que modulan la actividad del propio canal (Vriens et al., 2014; Diver et al., 2019).
De hecho, TRPM8 presenta fenómenos de desensibilización dependientes de calcio, en los que el aumento de Ca²⁺ intracelular conduce a una reducción progresiva de la actividad del canal. Este proceso está relacionado, al menos en parte, con la regulación de PIP2 y con la activación de proteínas moduladoras que afectan la estabilidad del canal en su estado abierto (Rohács et al., 2005).
Además, TRPM8 puede ser modulado por cambios en el potencial de membrana, mostrando cierta dependencia voltaje-sensible que, aunque no es dominante, contribuye a ajustar su actividad en función del estado eléctrico de la célula. Esta propiedad añade una capa adicional de integración de señales en su funcionamiento (Voets et al., 2004).
En conjunto, los mecanismos moleculares de TRPM8 reflejan un sistema altamente integrado en el que temperatura, ligandos químicos, lípidos de membrana y señales intracelulares convergen para regular la apertura del canal. Esta integración permite una detección precisa y adaptable de estímulos fríos en distintos contextos fisiológicos (Yin et al., 2018; Vriens et al., 2014).
Farmacología
La farmacología de TRPM8 se caracteriza por la existencia de múltiples agonistas y antagonistas que modulan su actividad mediante interacción directa con el canal o a través de cambios en su entorno lipídico. Estos compuestos han sido fundamentales tanto para su caracterización funcional como para explorar su potencial como diana farmacológica (De Petrocellis et al., 2011).
Entre los agonistas más conocidos se encuentran compuestos que inducen sensación de frío, siendo el mentol el prototipo clásico. El mentol activa TRPM8 mediante unión a sitios específicos en el dominio transmembrana, provocando la apertura del canal y generando la típica sensación refrescante (McKemy et al., 2002; Bandell et al., 2004).
Otros agonistas incluyen moléculas sintéticas como icilina, que presenta una potente capacidad de activación del canal, aunque su mecanismo de acción difiere parcialmente del mentol y puede depender de condiciones intracelulares específicas, como la concentración de calcio (Behrendt et al., 2004).
En el ámbito de los antagonistas, diversos compuestos han sido desarrollados para inhibir la actividad de TRPM8, incluyendo moléculas sintéticas que bloquean el canal de manera directa. Estos antagonistas han sido utilizados principalmente en investigación para estudiar la función del canal en la percepción del frío y en modelos experimentales de dolor (Knowlton et al., 2011).
En relación con los cannabinoides, varios fitocannabinoides han demostrado capacidad para modular TRPM8, generalmente ejerciendo efectos inhibitorios. El cannabidiol (CBD), en particular, ha mostrado reducir la actividad del canal en sistemas experimentales, lo que refuerza su papel como modulador de dianas no clásicas del sistema endocannabinoide (De Petrocellis et al., 2011; Muller et al., 2019).
El Δ9-tetrahidrocannabinol (THC) y otros fitocannabinoides también pueden interactuar con canales TRP, aunque su efecto sobre TRPM8 es más variable y depende del contexto experimental, incluyendo concentración y sistema celular utilizado (De Petrocellis et al., 2011).
Además de ligandos directos, la actividad de TRPM8 puede verse modulada por factores indirectos como la composición lipídica de la membrana, la disponibilidad de PIP2 y la activación de rutas de señalización intracelular. Estos factores influyen en la sensibilidad del canal y en su respuesta a agonistas y antagonistas (Rohács et al., 2005).
Desde una perspectiva aplicada, TRPM8 ha sido considerado una posible diana en el desarrollo de compuestos destinados a modular la percepción sensorial, especialmente en contextos relacionados con el frío y la activación de fibras sensoriales periféricas. No obstante, su complejidad funcional y su amplia distribución plantean retos en la selectividad de posibles intervenciones farmacológicas (Vriens et al., 2014).
En conjunto, la farmacología de TRPM8 refleja la versatilidad de este canal como integrador de señales químicas y físicas, así como su potencial como diana en la modulación de la señalización sensorial (De Petrocellis et al., 2011).
Rol fisiológico
TRPM8 desempeña un papel central en la detección y codificación de estímulos térmicos fríos dentro del sistema somatosensorial, actuando como uno de los principales sensores periféricos responsables de traducir cambios de temperatura en señales eléctricas interpretables por el sistema nervioso central. Su activación en terminaciones nerviosas periféricas permite generar la percepción consciente del frío y desencadenar respuestas adaptativas frente a variaciones térmicas del entorno (Dhaka et al., 2007; Vriens et al., 2014).
En condiciones fisiológicas, TRPM8 se activa en rangos de temperatura moderadamente fríos, contribuyendo a la detección de estímulos no nocivos. Esta información es transmitida a través de fibras aferentes primarias hacia la médula espinal y estructuras superiores, donde se integra con otras señales sensoriales para generar una representación precisa del entorno térmico (Knowlton et al., 2010; Dhaka et al., 2006).
Además de su función en la termorrecepción, TRPM8 participa en la modulación de la señalización nociceptiva. Aunque tradicionalmente se ha asociado con la detección de frío inocuo, su activación en determinados contextos puede influir en la percepción del dolor, modulando la excitabilidad de circuitos neuronales implicados en la nocicepción. Esta dualidad funcional refleja la complejidad de su papel dentro del sistema sensorial (Knowlton et al., 2011; Vriens et al., 2014).
En este sentido, la activación de TRPM8 por compuestos como el mentol puede generar efectos de modulación sensorial que alteran la percepción de estímulos nocivos, lo que ha sido interpretado como un fenómeno de interacción entre vías sensoriales térmicas y nociceptivas. Estos efectos no implican necesariamente una acción directa sobre el dolor, sino una reorganización de la señal sensorial a nivel periférico y central (Bautista et al., 2007).
TRPM8 también está implicado en fenómenos de hipersensibilidad al frío en contextos patológicos. En condiciones como inflamación o daño nervioso, se ha observado un aumento en la actividad o sensibilidad de neuronas que expresan TRPM8, lo que puede dar lugar a respuestas exageradas frente a estímulos fríos (alodinia al frío). Este fenómeno pone de manifiesto su relevancia en la plasticidad del sistema sensorial (Vriens et al., 2014).
Más allá del sistema nervioso, la expresión de TRPM8 en tejidos no neuronales sugiere funciones adicionales en fisiología periférica. En el tracto respiratorio y en epitelios, su activación por estímulos fríos o químicos puede participar en respuestas locales, como la regulación del tono tisular o la percepción de flujo de aire, aunque estos roles no están completamente definidos (Bidaux et al., 2007).
En el ámbito urogenital, especialmente en próstata, TRPM8 ha sido estudiado por su expresión diferencial y su posible implicación en procesos celulares relacionados con la homeostasis tisular. Aunque estos hallazgos han generado interés en investigación biomédica, su función fisiológica exacta en estos tejidos sigue siendo objeto de estudio (Tsavaler et al., 2001; Bidaux et al., 2007).
Desde una perspectiva integradora, TRPM8 actúa como un nodo dentro de una red sensorial compleja en la que convergen estímulos térmicos, químicos y moduladores lipídicos. Su capacidad para integrar estas señales permite ajustar la percepción sensorial en función del contexto fisiológico, contribuyendo a la adaptación del organismo a su entorno (Vriens et al., 2014).
En conjunto, el rol fisiológico de TRPM8 trasciende la simple detección del frío, participando en la modulación de la señal sensorial y en procesos adaptativos que reflejan la complejidad del sistema somatosensorial. Su estudio continúa ampliando la comprensión de cómo los organismos interpretan y responden a estímulos ambientales (Dhaka et al., 2007).
Importancia en investigación científica
El estudio de TRPM8 ha contribuido de manera significativa a la comprensión molecular de la termorrecepción, consolidando la idea de que la percepción de la temperatura no es un fenómeno difuso, sino el resultado de la activación de sensores específicos altamente especializados. Su identificación permitió establecer un marco conceptual en el que distintos canales TRP actúan como detectores de rangos térmicos concretos, redefiniendo la base molecular de la percepción sensorial (McKemy et al., 2002; Dhaka et al., 2006).
Desde una perspectiva neurobiológica, TRPM8 ha servido como modelo para estudiar cómo los estímulos físicos, como la temperatura, pueden traducirse en señales eléctricas mediante cambios conformacionales en proteínas de membrana. Este enfoque ha ampliado la comprensión de los mecanismos de transducción sensorial y ha facilitado la identificación de principios generales aplicables a otros sistemas sensoriales (Vriens et al., 2014).
En el ámbito de la farmacología, TRPM8 ha emergido como una diana molecular relevante para investigar la modulación de la señalización sensorial. El uso de agonistas y antagonistas selectivos ha permitido desentrañar su función en distintos contextos fisiológicos, así como explorar su papel dentro de redes más amplias de señalización que incluyen canales TRP relacionados y mediadores lipídicos (De Petrocellis et al., 2011).
Además, el estudio de TRPM8 ha sido clave para comprender la interacción entre canales iónicos y el entorno lipídico de la membrana. Su dependencia de moléculas como PIP2 ha puesto de manifiesto la importancia de los lípidos en la regulación de proteínas de membrana, contribuyendo a una visión más integrada de la señalización celular (Rohács et al., 2005).
En el contexto del sistema endocannabinoide ampliado (endocannabinoidome), TRPM8 representa una diana molecular que ayuda a explicar la diversidad de efectos asociados a cannabinoides y otros lípidos bioactivos. Su modulación por estos compuestos ha reforzado la idea de que los efectos de estas moléculas no se limitan a los receptores CB1 y CB2, sino que involucran múltiples sistemas interconectados (Di Marzo & Piscitelli, 2015).
Asimismo, TRPM8 ha sido ampliamente utilizado en modelos experimentales para estudiar la percepción del frío y la plasticidad del sistema sensorial en condiciones fisiológicas y patológicas. Estos modelos han permitido analizar cómo cambios en la expresión o función del canal pueden alterar la percepción sensorial, proporcionando herramientas para investigar fenómenos como la hipersensibilidad al frío (Dhaka et al., 2007; Vriens et al., 2014).
Los avances en técnicas estructurales, especialmente mediante criomicroscopía electrónica, han convertido a TRPM8 en un sistema de referencia para estudiar la dinámica conformacional de canales iónicos. Estos estudios han permitido visualizar estados funcionales del canal y comprender con mayor precisión los mecanismos que subyacen a su activación (Yin et al., 2018; Diver et al., 2019).
En conjunto, TRPM8 ocupa una posición relevante en la investigación biomédica al situarse en la intersección entre neurobiología, biofísica, farmacología y señalización lipídica. Su estudio no solo ha ampliado el conocimiento sobre la percepción del frío, sino que también ha contribuido a desarrollar un marco más amplio para entender cómo los organismos integran estímulos físicos y químicos a nivel molecular (Vriens et al., 2014).
Referencias
- McKemy DD, Neuhausser WM, Julius D. (2002). Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature, 416(6876), 52–58. - Peier AM, Moqrich A, Hergarden AC, Reeve AJ, Andersson DA, Story GM, Earley TJ, Dragoni I, McIntyre P, Bevan S, Patapoutian A. (2002). A TRP channel that senses cold stimuli and menthol. Cell, 108(5), 705–715. - Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D. (1997). The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature, 389(6653), 816–824. - Dhaka A, Viswanath V, Patapoutian A. (2006). TRP ion channels and temperature sensation. Annual Review of Neuroscience, 29, 135–161. - Dhaka A, Murray AN, Mathur J, Earley TJ, Petrus MJ, Patapoutian A. (2007). TRPM8 is required for cold sensation in mice. Neuron, 54(3), 371–378. - Bautista DM, Siemens J, Glazer JM, Tsuruda PR, Basbaum AI, Stucky CL, Jordt SE, Julius D. (2007). The menthol receptor TRPM8 is the principal detector of environmental cold. Nature, 448(7150), 204–208. - Knowlton WM, Daniels RL, Palkar R, McCoy DD, McKemy DD. (2010). Pharmacological blockade of TRPM8 ion channels alters cold and cold pain responses in mice. PLoS One, 5(9), e13273. - Knowlton WM, Palkar R, Lippoldt EK, McCoy DD, Baluch F, Chen J, McKemy DD. (2011). A sensory-labeled line for cold: TRPM8-expressing sensory neurons define the cellular basis for cold, cold pain, and cooling-mediated analgesia. Journal of Neuroscience, 31(8), 2837–2848. - Bandell M, Story GM, Hwang SW, Viswanath V, Eid SR, Petrus MJ, Earley TJ, Patapoutian A. (2004). Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron, 41(6), 849–857. - Voets T, Droogmans G, Wissenbach U, Janssens A, Flockerzi V, Nilius B. (2004). The principle of temperature-dependent gating in cold- and heat-sensitive TRP channels. Nature, 430(7001), 748–754. - Rohács T, Lopes CM, Michailidis I, Logothetis DE. (2005). PI(4,5)P2 regulates the activation and desensitization of TRPM8 channels through the TRP domain. Nature Neuroscience, 8(5), 626–634. - Liu B, Qin F. (2005). Functional control of cold- and menthol-sensitive TRPM8 ion channels by phosphatidylinositol 4,5-bisphosphate. Journal of Neuroscience, 25(7), 1674–1681. - Tsavaler L, Shapero MH, Morkowski S, Laus R. (2001). Trp-p8, a novel prostate-specific gene, is up-regulated in prostate cancer and other malignancies and shares high homology with transient receptor potential calcium channel proteins. Cancer Research, 61(9), 3760–3769. - Bidaux G, Roudbaraki M, Merle C, Crépin A, Delcourt P, Slomianny C, Thebault S, Bonnal JL, Benahmed M, Cabon F, Mauroy B, Prevarskaya N. (2007). Evidence for specific TRPM8 expression in human prostate secretory epithelial cells: functional androgen receptor requirement. Endocrine-Related Cancer, 14(3), 701–714. - De Petrocellis L, Orlando P, Moriello AS, Aviello G, Stott C, Izzo AA, Di Marzo V. (2011). Cannabinoid actions at TRP channels: effects on TRPM8 and TRPA1. British Journal of Pharmacology, 163(7), 1479–1494. - Müller C, Morales P, Reggio PH. (2019). Cannabinoid ligands targeting TRP channels. Frontiers in Molecular Neuroscience, 11, 487. - Di Marzo V, De Petrocellis L, Fezza F, Ligresti A, Bisogno T. (2002). Anandamide receptors. Prostaglandins, Leukotrienes and Essential Fatty Acids, 66(2–3), 377–391. - Di Marzo V, Piscitelli F. (2015). The endocannabinoid system and its modulation by phytocannabinoids. Neurotherapeutics, 12(4), 692–698. - Vriens J, Nilius B, Voets T. (2014). Peripheral thermosensation in mammals. Nature Reviews Neuroscience, 15(9), 573–589. - Yin Y, Wu M, Zubcevic L, Borschel WF, Lander GC, Lee SY. (2018). Structure of the cold- and menthol-sensing ion channel TRPM8. Science, 359(6372), 237–241. - Diver MM, Cheng Y, Julius D. (2019). Structural insights into TRPM8 inhibition and desensitization. Science, 365(6460), 1434–1440.