Introducción
El 2-arachidonoylglycerol (2-AG) es uno de los principales endocannabinoides descritos hasta la fecha y constituye, junto con la anandamida (AEA), uno de los ligandos endógenos más relevantes del sistema endocannabinoide. Se trata de un mediador lipídico derivado del ácido araquidónico que participa en múltiples procesos de señalización celular y desempeña un papel central en la regulación de la homeostasis biológica. A diferencia de neurotransmisores clásicos almacenados en vesículas, el 2-AG se sintetiza “a demanda” a partir de precursores lipídicos de membrana y actúa de forma transitoria en contextos fisiológicos específicos, especialmente en el sistema nervioso y en diversos tejidos periféricos (Kano et al., 2009; Di Marzo, 2018).
Desde el punto de vista bioquímico, el 2-AG pertenece a la familia de los monoacilglicéridos y presenta afinidad funcional por los receptores cannabinoides CB1 y CB2, sobre los que actúa como agonista endógeno. Su relevancia biológica ha sido ampliamente estudiada en neurobiología, farmacología y fisiología molecular, ya que interviene en fenómenos como la modulación sináptica retrógrada, la comunicación intercelular dependiente de lípidos y la regulación de respuestas inmunológicas y metabólicas. En comparación con la anandamida, el 2-AG suele encontrarse en concentraciones tisulares más elevadas y se considera un componente esencial de la señalización cannabinoide basal y actividad-dependiente (Sugiura et al., 1995; Stella et al., 1997).
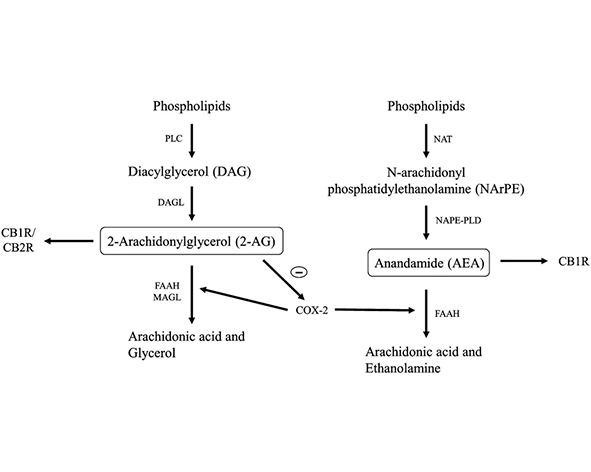
El interés científico por el 2-AG aumentó de forma notable a partir de la década de 1990, cuando distintos grupos de investigación comenzaron a identificar no solo su presencia endógena en mamíferos, sino también su papel como ligando fisiológicamente activo de los receptores cannabinoides. Desde entonces, su estudio ha permitido comprender con mayor precisión cómo el sistema endocannabinoide participa en la regulación de funciones como la neurotransmisión, la inflamación, el metabolismo energético y la plasticidad sináptica. Además, el análisis de sus rutas de síntesis y degradación —principalmente mediadas por enzimas como DAGL y MAGL— ha contribuido a consolidar una visión más completa del sistema endocannabinoide como red de señalización dinámica y finamente regulada (Mechoulam et al., 1995; Blankman et al., 2007).
En el contexto del sistema endocannabinoide, el 2-AG no debe entenderse como una molécula aislada, sino como parte de una arquitectura fisiológica compleja que integra receptores, enzimas, transportadores y señales intracelulares. Su estudio resulta fundamental para comprender tanto la biología básica del sistema como las preguntas abiertas que siguen marcando la investigación contemporánea en cannabinoides endógenos. Por ello, el 2-AG ocupa una posición central dentro de cualquier revisión científica dedicada a los fundamentos moleculares del sistema endocannabinoide (Di Marzo & Piscitelli, 2015; Lu & Mackie, 2016).
Historia del descubrimiento
El 2-arachidonoylglycerol (2-AG) fue identificado a mediados de la década de 1990 como parte de los esfuerzos por caracterizar ligandos endógenos capaces de activar los receptores cannabinoides previamente descritos. Tras el descubrimiento de la anandamida (AEA) en 1992, la investigación en el campo de los endocannabinoides se intensificó, dando lugar a la identificación de nuevas moléculas lipídicas con actividad biológica similar. En este contexto, el 2-AG fue descrito de manera independiente por varios grupos de investigación en 1995, consolidándose rápidamente como un componente clave del sistema endocannabinoide (Mechoulam et al., 1995; Sugiura et al., 1995).
Los primeros estudios demostraron que el 2-AG no solo estaba presente en tejidos de mamíferos, sino que además actuaba como un agonista completo de los receptores cannabinoides CB1 y CB2, diferenciándose en este aspecto de la anandamida, cuya actividad funcional es más compleja. Este hallazgo fue relevante, ya que indicaba que el sistema endocannabinoide no dependía de un único ligando endógeno, sino de un conjunto de moléculas con perfiles farmacológicos distintos. A partir de estos trabajos iniciales, el 2-AG comenzó a considerarse un mediador central en la señalización cannabinoide fisiológica (Sugiura et al., 1995; Stella et al., 1997).
Durante los años posteriores, la investigación se centró en determinar las rutas metabólicas implicadas en la síntesis y degradación del 2-AG, así como su distribución en distintos tejidos. Estos estudios permitieron identificar enzimas clave como la diacilglicerol lipasa (DAGL) en su síntesis y la monoacilglicerol lipasa (MAGL) en su degradación, estableciendo así el marco bioquímico básico que define su dinámica en el organismo. Este avance contribuyó a consolidar la idea del sistema endocannabinoide como una red de señalización lipídica altamente regulada y dependiente del contexto fisiológico (Bisogno et al., 2003; Blankman et al., 2007).
Con el desarrollo de nuevas técnicas analíticas y modelos experimentales, el 2-AG pasó de ser considerado un simple ligando endógeno a un elemento central en la comprensión de procesos como la plasticidad sináptica, la señalización retrógrada y la modulación de circuitos neuronales. En particular, estudios en neurobiología demostraron que el 2-AG actúa como mediador clave en la supresión transitoria de la liberación de neurotransmisores, lo que ha reforzado su papel en la regulación dinámica de la comunicación neuronal. Estos descubrimientos han situado al 2-AG en el núcleo de la investigación moderna sobre el sistema endocannabinoide (Kano et al., 2009; Castillo et al., 2012).
En la actualidad, el 2-AG es ampliamente reconocido como uno de los principales endocannabinoides tanto por su abundancia como por su relevancia funcional. Su descubrimiento y caracterización han sido fundamentales para comprender que el sistema endocannabinoide no es un sistema lineal, sino una red compleja en la que múltiples ligandos, enzimas y receptores interactúan de manera coordinada. Este cambio de paradigma ha marcado el desarrollo del campo y continúa guiando la investigación en fisiología, farmacología y biología molecular de los cannabinoides (Di Marzo & Piscitelli, 2015; Lu & Mackie, 2016).
Estructura o componentes
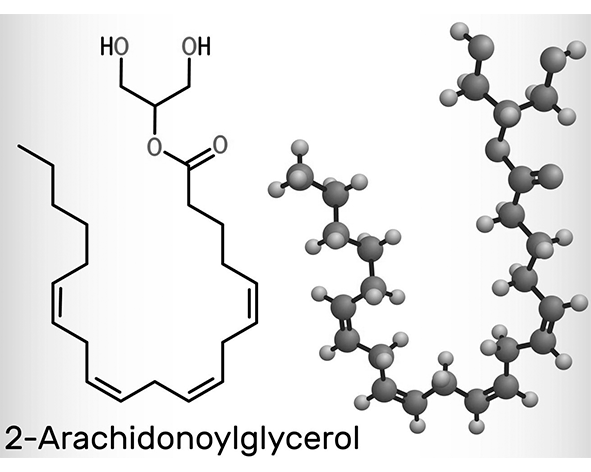
El 2-arachidonoylglycerol (2-AG) es un lípido perteneciente a la familia de los monoacilglicéridos, caracterizado por la esterificación del ácido araquidónico en la posición sn-2 del glicerol. Desde el punto de vista estructural, se trata de una molécula anfipática que combina una cadena hidrocarbonada poliinsaturada con un esqueleto de glicerol, lo que le confiere propiedades físico-químicas adecuadas para interactuar con membranas celulares y proteínas de señalización. Esta naturaleza lipídica condiciona su síntesis, transporte y degradación, diferenciándolo de neurotransmisores hidrosolubles clásicos (Sugiura et al., 1995; Piomelli, 2003).
El ácido araquidónico, componente central del 2-AG, es un ácido graso poliinsaturado de 20 carbonos (20:4, ω-6) que actúa como precursor de múltiples mediadores lipídicos. Su incorporación en la estructura del 2-AG permite que esta molécula participe en rutas metabólicas compartidas con otros sistemas de señalización, como los eicosanoides, estableciendo conexiones entre el sistema endocannabinoide y otras redes bioquímicas implicadas en la regulación fisiológica. Esta interconexión resalta el papel del 2-AG como nodo integrador dentro de la señalización lipídica celular (Di Marzo et al., 1998; Ahn et al., 2008).
La síntesis del 2-AG se produce principalmente a partir de diacilglicéridos (DAG) derivados de fosfolípidos de membrana, mediante la acción de enzimas específicas conocidas como diacilglicerol lipasas (DAGL), especialmente las isoformas DAGLα y DAGLβ. Este proceso ocurre de forma dependiente de la actividad celular, lo que significa que el 2-AG no se almacena en vesículas, sino que se genera “a demanda” en respuesta a estímulos fisiológicos. Este mecanismo de síntesis dinámica es una característica definitoria de los endocannabinoides y permite una regulación espacial y temporal precisa de la señalización (Bisogno et al., 2003; Gao et al., 2010).
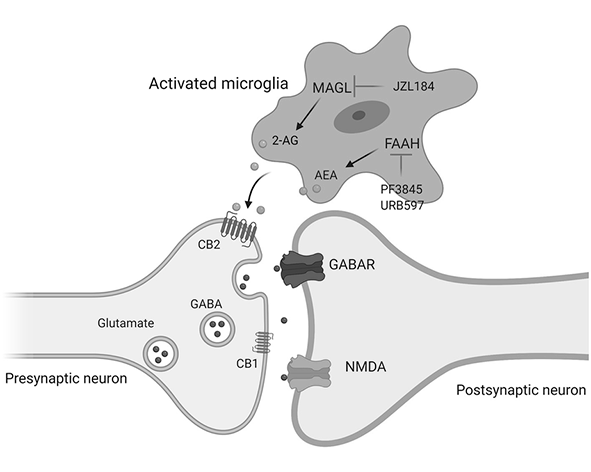
En cuanto a su degradación, el 2-AG es hidrolizado principalmente por la monoacilglicerol lipasa (MAGL), una enzima responsable de la mayor parte de su catabolismo en el sistema nervioso y otros tejidos. Adicionalmente, otras enzimas como ABHD6 y ABHD12 contribuyen a su degradación en compartimentos celulares específicos, modulando así la duración y magnitud de su señalización. Este equilibrio entre síntesis y degradación es fundamental para mantener niveles fisiológicos adecuados de 2-AG y evitar una activación desregulada de los receptores cannabinoides (Blankman et al., 2007; Savinainen et al., 2012).
Desde una perspectiva funcional, la estructura del 2-AG le permite actuar como un ligando lipofílico altamente móvil dentro del entorno de la membrana, facilitando su difusión lateral y su interacción con receptores acoplados a proteína G como CB1 y CB2. Esta capacidad de interacción directa con componentes de la membrana celular es clave para su papel en la señalización retrógrada y en la modulación de la transmisión sináptica. En conjunto, la organización estructural y metabólica del 2-AG refleja una adaptación evolutiva a la señalización rápida, localizada y reversible en sistemas biológicos complejos (Kano et al., 2009; Lu & Mackie, 2016).
Mecanismos moleculares
El 2-arachidonoylglycerol (2-AG) actúa principalmente como un ligando endógeno de los receptores cannabinoides CB1y CB2, ambos pertenecientes a la familia de los receptores acoplados a proteína G (GPCR). Tras su liberación desde la membrana postsináptica, el 2-AG difunde a través del espacio extracelular y se une a estos receptores, activando cascadas de señalización intracelular mediadas principalmente por proteínas Gi/o. Esta activación conduce a la inhibición de la adenilato ciclasa, la reducción de los niveles de cAMP y la modulación de diversas rutas de señalización celular (Howlett et al., 2002; Pertwee, 2008).
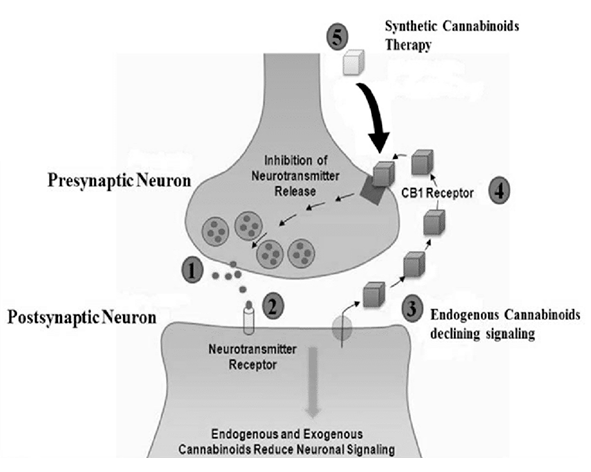
Uno de los mecanismos mejor caracterizados del 2-AG es la señalización retrógrada sináptica, un proceso mediante el cual las neuronas postsinápticas regulan la actividad de las neuronas presinápticas. En este contexto, el 2-AG se sintetiza en la neurona postsináptica en respuesta a un aumento de calcio intracelular o a la activación de receptores acoplados a Gq/11, y posteriormente viaja hacia la terminal presináptica, donde activa receptores CB1. Esta activación reduce la liberación de neurotransmisores como glutamato o GABA, modulando así la excitabilidad neuronal y contribuyendo a la regulación de la transmisión sináptica (Kano et al., 2009; Castillo et al., 2012).
A nivel intracelular, la activación de los receptores cannabinoides por 2-AG también influye en múltiples rutas de señalización, incluyendo las vías MAPK/ERK, PI3K/Akt y la regulación de canales iónicos dependientes de voltaje. Estas rutas participan en procesos como la supervivencia celular, la plasticidad sináptica y la adaptación a estímulos fisiológicos. Además, la activación de CB1 puede modular la apertura de canales de calcio tipo N y P/Q, así como la activación de canales de potasio, lo que contribuye a la disminución de la liberación de neurotransmisores en la sinapsis (Mackie, 2006; Turu & Hunyady, 2010).
El 2-AG también presenta un papel relevante en la señalización en células no neuronales, incluyendo células del sistema inmunitario, donde la activación de CB2 modula la liberación de mediadores inflamatorios y la migración celular. Aunque los mecanismos exactos pueden variar según el tipo celular y el contexto fisiológico, la activación de CB2 por 2-AG se asocia generalmente con la regulación de respuestas inmunológicas y la modulación de la señalización intracelular dependiente de quinasas y factores de transcripción (Turcotte et al., 2016; Cabral & Griffin-Thomas, 2009).
Es importante destacar que la señalización mediada por 2-AG está estrechamente regulada tanto por su síntesis localizada como por su rápida degradación, lo que limita su acción a escalas temporales cortas y evita una activación prolongada de los receptores cannabinoides. Este control dinámico permite que el 2-AG actúe como un modulador fino de la actividad celular, integrando señales eléctricas, químicas y metabólicas en diferentes sistemas fisiológicos. En conjunto, estos mecanismos posicionan al 2-AG como un mediador clave en la comunicación celular dependiente de lípidos dentro del sistema endocannabinoide (Piomelli, 2003; Lu & Mackie, 2016).
Distribución o contexto fisiológico
El 2-arachidonoylglycerol (2-AG) se encuentra ampliamente distribuido en el organismo, con presencia significativa tanto en el sistema nervioso central como en múltiples tejidos periféricos. En el cerebro, el 2-AG es uno de los endocannabinoides más abundantes y se localiza en regiones clave como el hipocampo, la corteza cerebral, el cerebelo y los ganglios basales, donde participa activamente en la regulación de la transmisión sináptica y la plasticidad neuronal. Esta distribución no es uniforme, sino que responde a la expresión diferencial de enzimas como DAGL y MAGL, así como a la densidad de receptores cannabinoides en cada región (Bisogno et al., 2003; Kano et al., 2009).
A nivel sináptico, el 2-AG se produce principalmente en neuronas postsinápticas y actúa sobre terminales presinápticas ricas en receptores CB1, configurando un sistema de comunicación retrógrada altamente localizado. Esta organización permite una regulación precisa de la liberación de neurotransmisores, contribuyendo al equilibrio entre excitación e inhibición en circuitos neuronales complejos. Además, el 2-AG se ha implicado en fenómenos como la depresión sináptica a corto y largo plazo, lo que refuerza su papel en la adaptación funcional del sistema nervioso (Castillo et al., 2012; Katona & Freund, 2012).
Fuera del sistema nervioso central, el 2-AG también se detecta en tejidos periféricos como el sistema inmunitario, el hígado, el tejido adiposo, el tracto gastrointestinal y el sistema cardiovascular. En células inmunitarias, incluyendo macrófagos y linfocitos, el 2-AG interactúa principalmente con receptores CB2, modulando procesos relacionados con la migración celular, la señalización intracelular y la liberación de mediadores inflamatorios. Esta distribución periférica sugiere que el sistema endocannabinoide no está restringido al sistema nervioso, sino que actúa como un sistema regulador global (Cabral & Griffin-Thomas, 2009; Turcotte et al., 2016).
En el contexto metabólico, el 2-AG se encuentra en órganos implicados en el metabolismo energético, como el hígado y el tejido adiposo, donde participa en la regulación de procesos relacionados con el almacenamiento y utilización de energía. Asimismo, su presencia en el tracto gastrointestinal y en el sistema endocrino indica un posible papel en la integración de señales hormonales y metabólicas, aunque los mecanismos específicos continúan siendo objeto de investigación. Esta distribución amplia refuerza la idea del sistema endocannabinoide como una red fisiológica transversal (Di Marzo & Matias, 2005; Silvestri & Di Marzo, 2013).
Es importante destacar que los niveles de 2-AG pueden variar en función del estado fisiológico, la actividad neuronal y las condiciones ambientales, lo que refleja su carácter dinámico como mediador lipídico. Factores como el estrés, la actividad física o cambios en la disponibilidad de nutrientes pueden influir en su síntesis y degradación, modulando así su acción en distintos tejidos. En conjunto, la distribución del 2-AG evidencia su papel como regulador sistémico que conecta múltiples sistemas fisiológicos a través de mecanismos de señalización dependientes de lípidos (Di Marzo, 2018; Lu & Mackie, 2016).
Interacción con fitocannabinoides
El 2-arachidonoylglycerol (2-AG) interactúa de forma indirecta con los fitocannabinoides, principalmente a través de la modulación compartida de los receptores cannabinoides CB1 y CB2, así como de las enzimas implicadas en su metabolismo. Aunque el 2-AG es un ligando endógeno y los fitocannabinoides son compuestos exógenos derivados de la planta Cannabis sativa, ambos convergen en los mismos sistemas de señalización, generando interacciones funcionales que dependen del contexto fisiológico y de la concentración relativa de cada molécula (Pertwee, 2008; Lu & Mackie, 2016).
El Δ9-tetrahidrocannabinol (THC), principal fitocannabinoide psicoactivo, actúa como agonista parcial de los receptores CB1 y CB2, compartiendo diana molecular con el 2-AG. Esta coincidencia implica que el THC puede competir con el 2-AG por la activación de estos receptores, alterando la señalización endocannabinoide basal. Sin embargo, a diferencia del 2-AG, cuya síntesis y liberación están finamente reguladas en tiempo y espacio, el THC introduce una activación más sostenida y menos controlada del sistema, lo que modifica la dinámica fisiológica de la señalización cannabinoide (Howlett et al., 2002; Pertwee, 2008).
Por otro lado, el cannabidiol (CBD) presenta un perfil farmacológico distinto, ya que no actúa como agonista directo de CB1 o CB2, sino que modula el sistema endocannabinoide a través de múltiples mecanismos indirectos. Entre ellos, se ha descrito su capacidad para influir en la degradación de endocannabinoides, incluyendo la inhibición parcial de enzimas como FAAH, lo que puede alterar los niveles de ligandos endógenos como la anandamida. Aunque el efecto del CBD sobre el metabolismo específico del 2-AG es más complejo y menos directo, su acción moduladora contribuye a modificar el equilibrio global del sistema endocannabinoide (Bisogno et al., 2001; Ibeas Bih et al., 2015).
Además de THC y CBD, otros fitocannabinoides y compuestos presentes en el cannabis, como los terpenos, pueden influir en la señalización del 2-AG a través de mecanismos complementarios, incluyendo la modulación alostérica de receptores o la interacción con otras dianas moleculares como canales TRP y receptores nucleares. Estas interacciones forman parte de lo que se ha denominado “efecto séquito” (entourage effect), un concepto que describe la acción combinada de múltiples compuestos sobre el sistema endocannabinoide, aunque su base mecanística continúa siendo objeto de investigación (Russo, 2011; Morales et al., 2017).
En conjunto, la relación entre el 2-AG y los fitocannabinoides no debe entenderse como una sustitución funcional, sino como una superposición de señales que pueden modificar la actividad del sistema endocannabinoide en diferentes niveles. Mientras que el 2-AG actúa como mediador fisiológico endógeno, los fitocannabinoides introducen variables externas que pueden alterar su señalización, ofreciendo un marco complejo para el estudio de la interacción entre compuestos endógenos y exógenos en sistemas biológicos (Di Marzo & Piscitelli, 2015; Lu & Mackie, 2016).
Importancia en investigación biomédica
El 2-arachidonoylglycerol (2-AG) ha adquirido una relevancia creciente en la investigación biomédica debido a su papel central dentro del sistema endocannabinoide y a su implicación en múltiples procesos fisiológicos fundamentales. Su abundancia relativa en tejidos, especialmente en el sistema nervioso, y su capacidad para activar de forma eficaz los receptores CB1 y CB2 lo posicionan como uno de los principales mediadores de la señalización cannabinoide endógena. Por ello, el 2-AG se ha convertido en una molécula de referencia en estudios destinados a comprender la regulación de la comunicación celular dependiente de lípidos (Kano et al., 2009; Lu & Mackie, 2016).
En el ámbito de la neurobiología, el 2-AG ha sido ampliamente investigado por su papel en la modulación sináptica y en procesos de plasticidad neuronal, incluyendo fenómenos como la depresión sináptica a corto y largo plazo. Modelos experimentales han permitido observar cómo la señalización mediada por 2-AG contribuye a la regulación dinámica de circuitos neuronales, integrando señales eléctricas y bioquímicas en el sistema nervioso central. Estos hallazgos han sido clave para entender mecanismos fundamentales de la neurotransmisión y la adaptación neuronal (Castillo et al., 2012; Katona & Freund, 2012).
En sistemas periféricos, el 2-AG también ha sido objeto de estudio en relación con la regulación inmunológica y el metabolismo energético. Investigaciones en modelos celulares y animales han explorado su papel en la modulación de la actividad de células inmunitarias, así como su participación en procesos metabólicos en tejidos como el hígado y el tejido adiposo. Aunque estos estudios han ampliado la comprensión del alcance fisiológico del sistema endocannabinoide, muchas de estas funciones continúan en fase de investigación y requieren una caracterización más detallada en contextos específicos (Cabral & Griffin-Thomas, 2009; Turcotte et al., 2016).
Otro aspecto relevante en la investigación biomédica del 2-AG es el estudio de las enzimas implicadas en su metabolismo, especialmente MAGL, ABHD6 y ABHD12, cuya modulación permite alterar los niveles endógenos de este ligando. El desarrollo de inhibidores selectivos de estas enzimas ha proporcionado herramientas experimentales para analizar la función del 2-AG en distintos sistemas biológicos, contribuyendo a una mejor comprensión de su papel en la señalización intracelular y en la homeostasis fisiológica (Blankman et al., 2007; Savinainen et al., 2012).
En conjunto, el estudio del 2-AG ha permitido avanzar en la caracterización del sistema endocannabinoide como una red compleja y altamente regulada, implicada en múltiples niveles de organización biológica. Más allá de sus funciones específicas, el 2-AG representa un modelo de cómo los mediadores lipídicos pueden actuar como señales dinámicas en sistemas biológicos, integrando procesos neuronales, inmunológicos y metabólicos. Por ello, continúa siendo un foco central en la investigación contemporánea en fisiología y farmacología de los cannabinoides (Di Marzo & Piscitelli, 2015; Iannotti et al., 2016).
Referencias
- Mechoulam R, Ben-Shabat S, Hanus L, Ligumsky M, Kaminski NE, Schatz AR, et al. (1995). Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochemical Pharmacology, 50(1), 83–90. - Sugiura T, Kondo S, Sukagawa A, Nakane S, Shinoda A, Itoh K, et al. (1995). 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochemical and Biophysical Research Communications, 215(1), 89–97. - Stella N, Schweitzer P, Piomelli D. (1997). A second endogenous cannabinoid that modulates long-term potentiation. Nature, 388(6644), 773–778. - Di Marzo V, Melck D, Bisogno T, De Petrocellis L. (1998). Endocannabinoids: endogenous cannabinoid receptor ligands with neuromodulatory action. Trends in Neurosciences, 21(12), 521–528. - Piomelli D. (2003). The molecular logic of endocannabinoid signalling. Nature Reviews Neuroscience, 4(11), 873–884. - Bisogno T, Howell F, Williams G, Minassi A, Cascio MG, Ligresti A, et al. (2003). Cloning of the first sn1-DAG lipases points to the spatial and temporal regulation of endocannabinoid signaling in the brain. Journal of Cell Biology, 163(3), 463–468. - Mackie K. (2006). Cannabinoid receptors as therapeutic targets. Annual Review of Pharmacology and Toxicology, 46, 101–122. - Blankman JL, Simon GM, Cravatt BF. (2007). A comprehensive profile of brain enzymes that hydrolyze the endocannabinoid 2-arachidonoylglycerol. Chemistry & Biology, 14(12), 1347–1356. - Kano M, Ohno-Shosaku T, Hashimotodani Y, Uchigashima M, Watanabe M. (2009). Endocannabinoid-mediated control of synaptic transmission. Physiological Reviews, 89(1), 309–380. - Katona I, Freund TF. (2012). Multiple functions of endocannabinoid signaling in the brain. Annual Review of Neuroscience, 35, 529–558. - Castillo PE, Younts TJ, Chávez AE, Hashimotodani Y. (2012). Endocannabinoid signaling and synaptic function. Neuron, 76(1), 70–81. - Ahn K, McKinney MK, Cravatt BF. (2008). Enzymatic pathways that regulate endocannabinoid signaling in the nervous system. Chemical Reviews, 108(5), 1687–1707. - Savinainen JR, Saario SM, Laitinen JT. (2012). The serine hydrolases MAGL, ABHD6 and ABHD12 as guardians of 2-arachidonoylglycerol signaling. Chemical Reviews, 112(4), 1826–1846. - Cabral GA, Griffin-Thomas L. (2009). Emerging role of the cannabinoid receptor CB2 in immune regulation. Expert Reviews in Molecular Medicine, 11, e3. - Turcotte C, Blanchet MR, Laviolette M, Flamand N. (2016). The CB2 receptor and its role as a regulator of inflammation. Cellular and Molecular Life Sciences, 73(23), 4449–4470. - Di Marzo V, Piscitelli F. (2015). The endocannabinoid system and its modulation by phytocannabinoids. Neurotherapeutics, 12(4), 692–698. - Lu HC, Mackie K. (2016). An introduction to the endogenous cannabinoid system. Biological Psychiatry, 79(7), 516–525. - Iannotti FA, Di Marzo V, Petrosino S. (2016). Endocannabinoids and endocannabinoid-related mediators: Targets, metabolism and role in neurological disorders. Progress in Lipid Research, 62, 107–128.